
Structural Bases of Ionotropic GABA Receptor Function and Modulation*
Yongchang Chang, MD, PhD
Division of Neurobiology, Barrow Neurological Institute, St. Joseph’s Hospital and Medical Center, Phoenix, Arizona
*Courtesy of Yongchang Chang, MD, PhD
The Chang Laboratory
Yongchang Chang received his MD in 1982 at Sun Yat-Sen University of Medical Sciences, Guangzhou, China and his PhD in 1998 at the University of Alabama at Birmingham. His early research experiences include studies of the role of endogenous opioid peptides in the pathogenesis of hemorrhagic shock and spectral analysis of the electroencephalogram. His current research interest is the structure-function relationship of GABA-gated ion channels.Yun Chen received her MD degree in 2002 at Zhengzhou University, and master’s degree in medical genetics in 2005 at Sun Yat-Sen University. She joined the laboratory in August 2005.
Neurotransmitter-operated (ligand-gated) ion channels play essential roles in communication between neurons. One major family of these ligand-gated ion channels includes nicotinic receptors, serotonin receptor type 3, GABA[A/C] receptors, and the glycine receptor. The research in our laboratory focuses primarily on the GABA receptors, the receptors for the major inhibitory neurotransmitter gamma-aminobutyric acid (GABA) in the brain. These receptors are the key targets of many clinically useful drugs for treatment of insomnia, anxiety, and epilepsy and for general anesthesia. The long-term goal of our laboratory is to provide structural bases for GABA receptor function and modulation and mechanisms of action for those drugs acting on GABA receptors by using an interdisciplinary approach.
Key Words: activation, allosteric modulation, barbiturates, benzodiazepines, desensitization, GABA receptor, luminescence resonance energy transfer, neurosteroids, photocrosslinking, site-specific flourescence, structure-function relationship
Abbreviations used: GABA, gamma aminobutyric acid; LRET, luminescence resonance energy transfer
The brain differs from other organs by its ability to process information via its interconnected neural network. The basic building blocks of the neural network, individual nerve cells called neurons, are excitable cells in the brain. Their excitability is controlled by a class of membrane proteins termed ion channels, which selectively allow charged particles (ions) to pass through the membrane and, in turn, control the membrane potential of neurons. Neurons can generate and propagate nerve pulses (action potentials) through voltage-gated (membrane potential-controlled) ion channels.
Communication between neurons, however, is mainly achieved by chemical signaling via neurotransmitters and their receptors (ligand-gated ion channels) at specialized sites termed synapses, where neurons form functional contacts with each other. To achieve full control, a neuron receives both excitatory and inhibitory synaptic inputs. Summation of these opposing synaptic inputs determines whether a neuron generates an action potential.
GABA is the major inhibitory neurotransmitter in the mammalian brain. As a neurotransmitter, GABA is released into a synaptic cleft by its presynaptic nerve terminus when a GABA-ergic (GABA releasing) neuron fires an action potential. Binding GABA to its receptors in the postsynaptic membrane mediates fast (through ionotropic receptors) and slow (through metabotropic receptors) synaptic inhibition on the postsynaptic neuron.
Based on their pharmacological properties, GABA receptors are classified into three types (A, B, and C). The GABA[B] receptor is a G-protein-linked receptor (metabotropic GABA receptor) and is not the interest of our study. The GABA[A] and GABA[C] receptors are GABA-gated chloride channels (ionotropic GABA receptors). On activation, the channel opens and allows an influx of negatively charged chloride ions through the pore. In turn, the postsynaptic membrane becomes hyperpolarized (resulting in a more negative intracellular potential) and produces an inhibitory effect on the postsynaptic neuron. Through this action, ionotropic GABA receptors play an important role in controlling neuronal excitability. The long-term goal of this laboratory is to provide structural bases for the mechanisms of the ionotropic GABA receptor function and mechanisms of action for drugs that act on ionotropic GABA receptors.
Structure and Function of Ionotropic GABA Receptors
Molecular cloning studies have revealed several different classes of the ionotropic GABA receptor subunits and their isoforms alpha 1-6, beta 1-3, gamma 1-3, delta, epsilon, rho 1-3, theta, pi, and chi. They are homologous with 70 to 80% amino acid identity among isoforms of each class and 30 to 40% sequence identity between classes.8 They all belong to a ligand-gated ion channel gene family, which includes nicotinic receptors, serotonin (5-HT3) receptor type 3, glycine receptors, and GABA[A] and GABA[C] receptors.[13,15]
The proposed topology for each subunit is a large extracellular N-terminal domain, four transmembrane domains (M1-M4), and a large intracellular loop between M3 and M4 (Fig. 1A). The extracellular N-terminal domain forms the agonist-binding pocket, presumably at the subunit-subunit interface. The ion-conducting pore is formed by transmembrane domains (M1-M4). The large intracellular loop can interact with intracellular proteins for receptor targeting or clustering.
Like other members of this receptor family, a functional GABA receptor has five subunits. The pentameric structure of the affinity purified native GABA[A] receptor was first directly demonstrated by electron microscopy (Fig. 1B).[11] A GABA receptor can be formed by five identical (homomeric) or different (heteromeric) subunits. Different subunits, isoforms, or their combinations can form functional channels with distinct properties, generating the functional diversity of GABA-gated ion channels. Theoretically, thousands of combinations of subunits and their isoforms are possible in a pentameric receptor. In reality, however, only a limited number of combinations exist. A typical GABA[A] receptor is composed of alpha, beta, and gamma subunits with a stoichiometry of two alpha, two beta‚ and one gamma subunits.[3]
The most abundant GABA[A] receptor subtype in the mammalian brain is alpha 1 beta 2 gamma 2.9 When exogenously expressed, the recombinant alpha 1 beta 2 gamma 2 GABA receptor has pharmacological and physiological properties similar to the typical native GABA[A] receptors, whereas the rho 1 subunit can form a homomeric GABA receptor with properties similar to native GABA[C] receptors.
receptor determined by electron microscopic image
analysis. J Neurochem 62(2):815-818, 1994, with permission from Blackwell Publishing.)“]
Structural Model of the N-Terminal Domain: GABA and Benzodiazepine Binding Sites
A typical GABA[A] receptor has two agonist binding pockets located in two subunit interfaces between beta and alpha subunits. The rho 1 homomeric GABA[C] receptor has five potential binding pockets in the subunit interfaces. The crystal structure of a homologous protein, acetylcholine binding protein, was used to generate homologous structural models of GABA[A] and GABA[C] receptor N-terminal domains (Fig. 2). The binding residues face subunit interfaces and form the putative binding pockets in the subunit interfaces. GABA[A] receptor activation also can be modulated by benzodiazepines. The benzodiazepine binding site is homologous to the agonist binding site, but it is located in a different subunit interface (between alpha 1 and gamma 2 subunits) in the N-terminal domain.
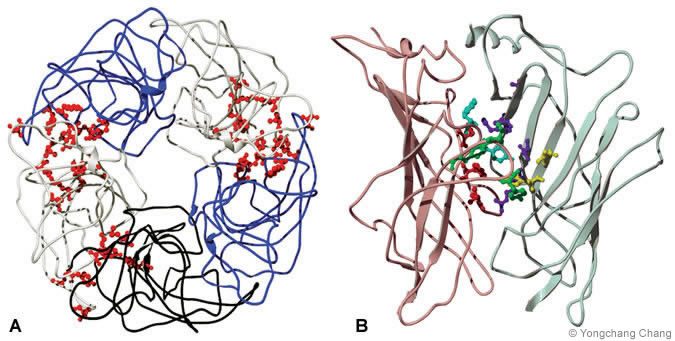{kind=link}
and (B) GABA[B] receptors.“]
Structural Model of Transmembrane Domains: Gating Machinery and Modulatory Sites
The ion channel part of ionotropic GABA receptors is formed by transmembrane domains. A structural model of GABA[A] receptor transmembrane domains was generated by homology modeling using the structure of the Torpedo nicotinic receptor based on electron microscopy (Fig. 3). Five subunits, each with four transmembrane domains (M1-M4), form a pseudosymmetric pentameric structure with the ion channel pore in the center. The second transmembrane domain (M2) lines the pore (Fig. 3A with M2 in the closed-channel position). The M2 domains are slightly kinked in the middle and form the ion channel gate to control opening and closing of the channel (Fig. 3B).
Accumulating evidence suggests that the transmembrane domains are also the major target for many clinically useful GABA[A] receptor modulators, such as neurosteroids, barbiturates, and general anesthetics (e.g., propofol).[12] Most of these compounds can potentiate the effect of GABA at low concentrations and directly activate the channel at high concentrations. Binding sites for these compounds are mainly located in the extracellular ends of M1, M2, and M3 (Fig. 3B). However, the detailed mechanisms of their action are still unknown.
receptor. The ion channel (center pore) is formed by five subunits, each with four transmembrane domains (M1-M4). (A) M2 lines the pore (top view). (B) Side view of transmembrane domains of one subunit with putative gate and binding sites (circle) for many GABA[A] receptor modulators.“]
GABA Receptor Activation: Coupling of Binding Domain to Channel Gate
Neurotransmitter-operated ion channels open in response to agonist binding. The conformational wave of the ion channel is initiated by the interaction of the agonist with its binding site. It must then propagate to the other parts of N-terminal domains and eventually to the gating machinery to open the channel.
A recent study with electron microscopy with a resolution of 4 Å has revealed more details of the activation mechanism of the Torpedo nicotinic receptor.10 In this model, the agonist binding to its binding pocket induces the N-terminal domain of each subunit to rotate. Subsequently, this rotation propagates to the gating machinery (M2) to open the pore through direct coupling between a short loop in the lower part of the N-terminal inner sheet and the end of M2.
In the heteromeric GABA[A] receptor, two agonist binding sites are available. Therefore, the receptor activation involves two binding steps,1 with the receptor at low affinity in the resting state(s) and at high affinity in the open state(s).[4] Although the detailed structural mechanism is unknown, channel opening involves conformational changes in both the gating machinery (opening) and the agonist binding pocket (increase in affinity). A recent study using site-directed mutagenesis and cross-linking suggested that the coupling mechanism for the channel activation is mediated by charge interactions between the N-terminal domain and M2-M3 linker (Fig. 4).[7] Activation of the homomeric rho 1 GABA[C] receptor requires three bindings to the five potential binding sites. Because gating can influence binding, channel opening of the homomeric rho 1 GABA[C] receptor can increase binding affinity to GABA to such an extent that the agonist appears to be locked in the binding site distant from the pore.[5] This increase in affinity during channel opening is similar to that in GABA[A] receptors. However, the detailed dynamic structural basis for this mutual influence of binding and gating during activation of GABA receptors is not yet established.
receptor is mediated by charge interaction between the N-terminal domain and M2-M3 linker (highlighted by circle).“]
GABA Receptor Desensitization: Uncoupling?
Binding of an agonist to the GABA[A], but not GABA[C], receptor can also simultaneously drive the receptor into a refractory state termed desensitization (decreased sensitivity). This state may play an important role in shaping synaptic inhibition. Desensitization of a ligand-gated ion channel involves conversion of the receptor from a low-affinity resting state to a high-affinity desensitized state2 and a change in the pore region to close the channel. Although the kinetic mechanism of desensitization for the GABA[A] receptor is well characterized,2 the structural basis of these changes remains poorly understood.
GABA[A] Receptor Allosteric Modulation: Drug Action
GABA[A] receptors can be potentiated by modulators, such as benzodiaz- epines, barbiturates, and neurosteroids. By binding to a site in the receptor distant from the GABA binding site, these modulators allosterically (distantly) modulate GABA[A] receptor function. Benzodiazepines are the best example of GABA[A] receptor modulators. Since their introduction into medicine in the 1960s, benzodiazepines are also among the most widely prescribed drugs in the world. Photoaffinity labeling and site-directed mutagenesis have identified residues for benzodiazepine binding in alpha (alpha 1H101, Y159, G200, T206, and Y209) and gamma (gamma 2 F77 and M130) subunits (Fig. 2A). Therefore, the benzodiazepine binding pocket is homologous to the GABA binding pocket but located in a different alpha-gamma subunit interface.[14] By binding to a site distinct from the GABA binding site, these compounds can change the apparent affinity of the GABA receptor for its agonist. However, the mechanism underlying this effect is poorly understood.
Since the early 1900s, barbiturates have been used as sedative-hypnotics, anticonvulsants, and anesthetics. They potentiate the GABA response of GABA[A] receptors at low concentrations and directly activate GABA[A] receptors at high concentrations. The site of action for barbiturates is not well defined, but it is distinct from the GABA binding site and involves the M1, M2 and M3 transmembrane domains. Recent studies with the substituted cysteine accessibility method have revealed that direct activation of the GABA[A] receptor by pentobarbital can cause the agonist binding pocket to narrow. Like barbiturates, neurosteroid anesthetics can potentiate the GABA effect at low concentrations and directly gate the GABA[A] receptor at high concentrations. The binding site for neurosteroids involves M2 and M4 transmembrane domains. However, experimental evidence suggests that they are distinct from barbiturate binding sites.
In summary, benzodiazepines, barbiturates, and neurosteroids have distinct binding sites, but they have similar (although not identical) potentiation effects on GABA[A] receptors. The structural basis for how these compounds have similar potentiation effects, by binding to distinct sites in a GABA[A] receptor, awaits future study.
Future Research
The current research interest of the laboratory is focused on the structural dynamics underlying GABA receptor function and modulation. Despite intensive structure-function relationship studies in the past, the dynamic structural basis underlying GABA receptor function and modulation is still poorly understood. To this end, the laboratory has successfully adapted several relatively new techniques to study the structural dynamics of the GABA receptor: site-specific fluorescence,[6] LRET, and photo crosslinking. The site-specific fluorescent technique allows the GABA-induced conformational change to be monitored at a specific site of the GABA receptor. The LRET technique makes it possible to measure distance changes, on an atomic scale, between two residues at different sites of the receptor during channel activation. The photo cross-linking technique immobilizes a specific moving part of the receptor, permitting the functional significance of the movement of a specific amino acid residue in the receptor to be evaluated. Using these newly established site-specific fluorescent, luminescent, and photochemical techniques, combined with electrophysiological, molecular biological, and computational modeling approaches, the laboratory has three immediate plans.
First, we will map the structural changes underlying the activation of the GABA[A/C] receptors. Although it is known that GABA binding induces conformational changes to open the channel, the detailed mechanism of this activation process is still unclear. We hypothesize that activation of the GABA receptor involves rotation of the receptor subunits, a mechanism similar to that proposed for the activation of a nicotinic receptor in the same gene family. Using our newly adapted techniques, we will systematically map the conformational changes in GABA[A/C] receptors during channel activation to gain insights into the detailed dynamic structural mechanism of GABA receptor activation.
Second, we hope to gain insights into the structural mechanism of GABA[A] receptor desensitization. Kinetically, the desensitization of GABA receptors is well characterized. Structurally, however, the process is poorly understood. We hypothesize that desensitization involves over-rotation of the N-terminal domain, which results in uncoupling of the N-terminus and gating machinery. The over-rotation also could change interacting residues in the N-terminal interface and stabilize the receptor in the high-affinity desensitized state. The newly adapted techniques will be used to systematically map the conformational rearrangements in GABA[A] receptors and to correlate these structural changes to functional desensitization to provide the dynamic structural basis of GABA-receptor desensitization.
Finally, we plan to define the structural basis underlying the modulation of GABA[A] receptors by benzodiazepines, barbiturates, and neurosteroids. As mentioned, these compounds bind to distinct sites of the GABA[A] receptor and potentiate the effect of GABA. By systematically mapping the conformational changes induced by these three different classes of compounds, we will be able to delineate the patterns of conformational changes underlying their allosteric modulation. This information will provide the dynamic structural basis of how the binding of these compounds to distinct sites can lead to similar potentiation actions. Systematic mapping of conformational changes should also suggest insights into the dynamic structural basis for alternative ways of channel activation by barbiturates and neurosteroids.
Significance
The ionotropic GABA receptors are widely distributed in the central nervous system and play a key role in brain function. Dysfunction of GABA receptors is implicated in the origin of a variety of neurological and psychiatric disorders such as epilepsy, anxiety, insomnia, and schizophrenia. GABA receptors are also the key targets for many widely prescribed neuroactive compounds, such as benzodiazepines, barbiturates, neurosteroids, and other general anesthetics. Therefore, it is paramount to elucidate the mechanism(s) underlying GABA receptor function and modulation.
Although in the past two decades intensive research has shaped a structural model of the ionotropic GABA receptors, the structural dynamics underlying their function and modulation are still not well established. Our work is intended to unveil novel dynamic structural mechanisms underlying activation, desensitization, and allosteric modulation of the GABA receptor. Gaining new insights into the structural dynamics underlying GABA receptor function and modulation is fundamental for understanding the pathogenesis of GABA receptor-related neurological and psychiatric disorders and the mechanism of action for those compounds with GABA receptors as their major targets. Such insights also facilitate the development of new drugs that can improve manipulation of GABA receptor function. In turn, new treatments may be generated for many neurological and psychiatric disorders, thereby improving quality of life. This research also will pave the way for future fluorescence-based studies, such as studying structural dynamics at the level of single molecules.
References
- Amin J, Weiss DS: GABA[A] receptor needs two homologous domains of the beta-subunit for activation by GABA but not by pentobarbital. Nature 366:565-569, 1993
- Chang Y, Ghansah E, Chen Y, et al: Desensitization mechanism of GABA receptors revealed by single oocyte binding and receptor function. J Neurosci 22:7982-7990, 2002
- Chang Y, Wang R, Barot S, et al: Stoichiometry of a recombinant GABA[A] receptor. J Neurosci 16:5415-5424, 1996
- Chang Y, Weiss DS: Allosteric activation mechanism of the alpha1beta2gamma2 gamma-aminobutyric acid type A receptor revealed by mutation of the conserved M2 leucine. Biophys J 77:2542-2551, 1999
- Chang Y, Weiss DS: Channel opening locks agonist onto the GABA receptor. Nat Neurosci 2:219-225, 1999
- Chang Y, Weiss DS: Site-specific fluorescence reveals distinct structural changes with GABA receptor activation and antagonism. Nat Neurosci 5:1163-1168, 2002
- Kash TL, Jenkins A, Kelley JC, et al: Coupling of agonist binding to channel gating in the GABA[A] receptor. Nature 421:272-275, 2003
- Macdonald RL, Olsen RW: GABA[A] receptor channels. Annu Rev Neurosci 17:569-602, 1994
- McKernan RM, Whiting PJ: Which GABA[A] receptor subtypes really occur in the brain? Trends Neurosci 19:139-143, 1996
- Miyazawa A, Fujiyoshi Y, Unwin N: Structure and gating mechanism of the acetylcholine receptor pore. Nature 423:949-955, 2003
- Nayeem N, Green TP, Martin IL, et al: Quaternary structure of the native GABA[A] receptor determined by electron microscopic image analysis. J Neurochem 62:815-818, 1994
- Olsen RW, Chang CS, Li G, et al: Fishing for allosteric sites on GABA(A) receptors. Biochem Pharmacol 68:1675-1684, 2004
- Ortells MO, Lunt GG: Evolutionary history of the ligand-gated ion-channel superfamily of receptors. Trends Neurosci 18:121-127, 1995
- Sawyer GW, Chiara DC, Olsen RW, et al: Identification of the bovine gamma-aminobutyric acid type A receptor alpha subunit residues photolabeled by the imidazobenzodiazepine [3H]Ro15-4513. J Biol Chem 277:50036-50045, 2002
- Unwin N: Nicotinic acetylcholine receptor at 9 Å resolution. J Mol Biol 229:1101-1124, 1993